熟人重要,还是族群重要? | Paper Alert

原创 NR 神经前研 收录于话题#Paper Alert25个
认知与行为
我们都一样——
人类大脑如何编码时空
Qasim et al., Cell
@图图
你知道从我们这里去茶颜悦色咋走吗?要多久?当问到这个问题时,你一定会头脑风暴一番,先判断自己在哪,走哪条路经过几个路口才能去到那个人满为患的奶茶店。那你的大脑是如何将这些信息联系起来?我们又为何记得何时何地发生过什么?
这些都要仰仗我们的大脑神经元活动有条不紊的工作。不同的神经元在不同地方不同时间有规划地活跃起来,编码着我们经历的顺序,能帮助我们的大脑形成一个“大局观”。特别的是,之前已有相关研究表明,啮齿类动物的海马体需要单个神经元通过相移(phase precession)有节奏的与局部θ振荡(∼5–10 Hz)合作实现对整体信息的整合压缩,以支撑我们对空间、时间等序列事件的认知、学习和记忆。但是,这十几年来,我们的研究都局限于现存的模式动物研究中,还并未在人体内发现这种相移的现象。
然而,近日,来自哥伦比亚大学工程与应用科学学院的Jacobs研究组,率先报道了他们在人体内的发现,人类大脑同样拥有着这种非常重要的时序编码功能,在空间探索过程中他们在人脑内嗅皮质(entorhinal cortex)和海马体中的神经元中,同样观测到了与啮齿类动物非常类似的相移。除了空间信息,人类大脑的相移还可以编码一些别的更为复杂的信息,比如它们与特殊的目标状态也息息相关。
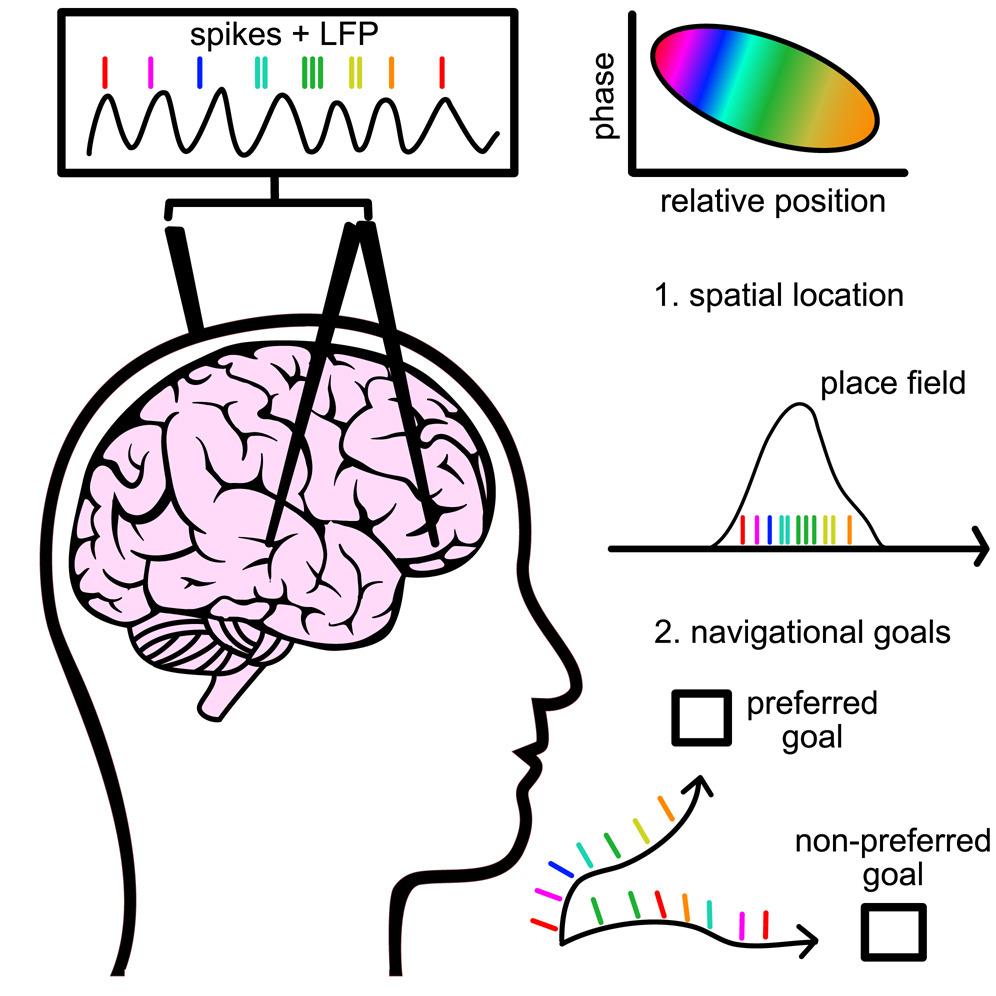 - Qasim et al., Cell -
- Qasim et al., Cell -为了记录并分析人类大脑中的神经元活动和电位等信息,研究人员分析了13个正在接受耐药性癫痫(drug-resistant epilepsy,DRE)临床治疗的并曾经接受过脑部神经手术(有植入电极)的病患不同脑区的神经元放电情况。这些受试者在虚拟现实空间导航任务中可以自由顺时针或逆时针探索。环境中包含有方形广场和周围的六座特定目标建筑物。
首先,研究人员比较了人类的啮齿类动物的海马体θ振荡,发现人类的θ振荡频率范围明显更广但低频尖峰值更低。在这个实验中,他们对相移的评估是基于2–10 Hz的θ振荡频率,之后,他们鉴别出了受试者海马体和内嗅皮质中与空间信息紧密联系的神经元和它们对应的放电尖峰位置(spike-phase),注意到它们刚开始都会在θ振荡波的相位后期放电。但随着移动时间的推移,它们的放电尖峰会有序的逐步向脑电波相位前移动。这说明神经元的放电尖峰时序与它们对应的空间位置呈负相关,从而根据神经元放电的尖峰相位可以推断出到达位置中心的路径中不同的相对位置。这与之前在啮齿动物中的观察结果非常类似。同时,他们注意到在空间探索任务中,人类大脑中感知位置信息的神经元其相移对空间位置更为敏感,而非单纯对时间流逝敏感,说明这些神经元放电相位能非常有效的编码空间位置信息。
但相移是否只与空间位置有关呢?因为之前在啮齿类动物中发现,还存在一些非空间类行为,例如快速动眼期或跳跃时编码时间等,表明大脑不同区域会普遍利用相移来编码系列事件顺序、状态。所以,人类是否也如此呢?接着研究人员通过测量神经元放电尖峰相位振谱进一步确认,人类大脑中非空间神经元也存在着非常统一有节律的放电现象(比θ振荡频率更高),也就是说这些神经元中也普遍存在着类似编码空间顺序的相移。那么这种非空间相移是否有可能在非空间性更复杂/高级的任务(比如寻找特定目标)中发生改变呢?它们是否有选择性的只在特殊目标相关轨迹中出现,从而有效的将这些轨迹信息结合在一起,进而服务于我们的序列学习和记忆?
在这一测试中,受试者需要根据给出的信号移动到随机选择的目标地点。研究人员发现,有些细胞在移动到一些特殊目标途中会非常有针对性的表现出相移,说明这些细胞(相对集中于前扣带皮质/眼窝前额皮质/杏仁核和海马体)的相移是仅与目标状态相关,而非其他可能的因素,例如不同目标本身引起不同的LFP功率或神经元放电速率,又或是受试者在任务中对不同目标的表现带来的差异,亦或是地理位置等。这些结果证明了,不仅在啮齿动物中,即使在人类大脑中,也存在着通过相移来编码非空间复杂事件的机制。而这极有可能是我们序列行为/学习的基础。
总的来说,这项研究将相移理论很严谨的延伸到了人类大脑,并拓展到了非空间顺序事件中,说明人类与之前的研究过的啮齿类动物模型非常类似。大脑中神经元与局部电位活动可以通过相移实现非常高效的合作共赢来编码时序,这是我们对事件产生有序认知和记忆的坚实神经基础。
doi: 10.1016/j.cell.2021.04.017
人脸识别的两个因素:
你熟悉那张脸吗?你们是同种族吗?
Zhou et al., Perception
@苏木弯
假如发生了一起事件,有两个目击证人声称看到了事件相关人。第一个目击证人与事件相关人熟悉,但不是同种族;第二个目击证人与事件相关人不熟,但属于同种族。谁的证词更值得相信呢?
人脸识别中一个著名的现象是“异族效应”(other-race effect),即同种族的脸比其他种族的脸更容易记忆。那么,把异族效应和其他因素一起考虑会怎么样呢?比如,熟悉度——“认识的其他人种的人”,和“不熟的同人种的人”,谁的脸更好记呢?在考虑是否有影响的基础上,影响有多大也是很关键的问题。Zhou等人通过实验和问题比较了熟悉度(熟悉/不熟悉)和种族(同种族/不同种族)这两个因素对人脸识别的影响的效果。他们的实验主要是一个新/旧识别测试。实验包括学习和测试两个阶段。在学习阶段,被试按随机顺序浏览64张不同的人脸图片(包括16个黑人名人,16个白人名人,16个黑人普通人,16个白人普通人)。在接下来的测试阶段,被试会按随机顺序看到128张人脸图片(包括32个黑人名人,32个白人名人,32个黑人普通人,32个白人普通人),其中每个类别都有一半(共64张)是在学习阶段出现过的旧脸,另一半是没有出现过的新脸。对于每一张图片,被试需要回答这张图片中的人脸是否在学习阶段出现过。
除了测量被试回答的准确性,实验还包括了一系列的问题以测量人们的“元认知”,也就是人们对于自己的认知的看法。问题分为自我评估和对他人评估。自我评估有三项:首先是在学习阶段的预期评估,对于每一张人脸图片,被试都要在百分比的度量上选择一个数来表示他们在记忆测试中记得这张脸的可能性;其次是在测试阶段的同步评估,对于每一个回答,被试都需要在百分比的度量上选择一个数来代表对于这个回答的自信程度;最后是测试结束后的调查问卷中的回顾评估,在学习阶段出现过的64张人脸会同时再次出现,并被分成2x2的表格(16个黑人名人,16个白人名人,16个黑人普通人,16个白人普通人),被试需要选择两个他们认为自己记得最好的分类。一共有六种选法。对他人评估包括在测试阶段的两个问题,对于每一张人脸图片,被试需要回答他们认为和这张人脸的主人同一种族以及不同种族的人的回答情况(分别在30个人中有多少人能回答正确)。最后被试需要填写他们自己的种族。
实验结果显示,种族和熟悉度都对人脸识别测试的表现有影响,但熟悉度的影响要强得多。效应值(effect size)的比较发现,熟悉度对于识别表现的影响比起种族至少要强10倍。由于在学习阶段和测试阶段使用了相同的图片,人脸识别表现可能归因于对特定的图片的记忆,而并不是单纯的人脸记忆。Zhou等人进行了第二次实验。在本次实验中,学习阶段和测试阶段使用了相同人脸的不同图片,以确保实验测试的是对人脸的记忆而不是对图片的记忆。两次实验的结果是相似的,证明第一次实验的结果并不是因为重复图片。
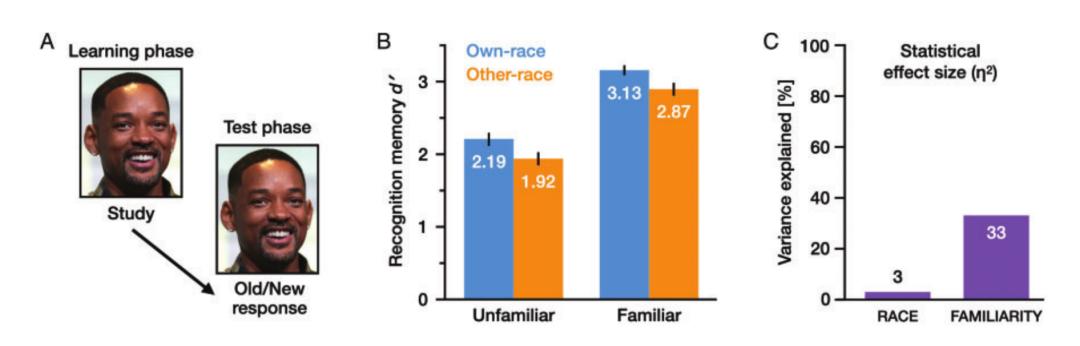 - Zhou et al., Perception -
- Zhou et al., Perception -自我评估的结果表示人们意识到了熟悉度的影响比种族的影响要大。在两次实验中,预期评估,同步评估,和实验后的回顾评估结果都和被试的真实表现相符。然而,对他人评估却显示出不同的结果。人们系统性地低估了他人的表现。另外,更重要的是,人们高估了异族效应对他人的影响,认为种族和熟悉度的影响相似。
在过去的实验以及社会生活中,我们都很关注种族对人脸识别记忆的影响。这种影响确实存在,但是这个影响是较小的,远不如熟悉度的影响。Zhou等人的实验中对于“元认知”的调查结果也显示,“异族效应”的社会重要性可能来源于人们对它认知的效果,而不是其真实效果。总结来说,Zhou等人的实验并不是要否定或者弱化“种族效应”,而是想要强调熟悉度的作用。所以,别担心,就算是其他种族的人的长相,脸熟之后也能记住!
doi: 10.1177/03010066211014016
我的舌头太固执,都是ALM的锅
Bollu et al., Nature
@图图
俗话说得好:“失败乃成功之母”,我们的行为是在一遍又一遍的尝试、错误然后修正中得以逐步完善的。比如,夜里摸黑找厕所,我们不可能第一次就非常顺利的摸到门把手,开门,找准位置,入坑(当然如果你有特异功能就另说)。正常人都需要慢慢摸索的吧?这种能够在尝试错误中更新动作信息以便快速准确找到目标的能力对我们的生存必不可少。
当然,不仅仅是我们的四肢动作需要精细调节,我们灵活的小舌头也有聪明的小眼睛,寻找美食、水源等不在话下。你有没有发现,就算闭着眼睛吃东西,第一次也许会磕到牙齿或者拍到脸上,但是两三次尝试后,就跟正常进食基本无异了,那是因为舌头可以准确的找到食物所在,进而暴风满足。在啮齿动物中,比如小鼠,“舔”(licking)出水口常常被用来研究决策选择、行动计划和实施等复杂的认知行为。但时至今日,我们的舌头是如何寻找到出水口的这一谜题还并没有得到很好的解释。近日,来自康奈尔大学的Goldberg实验室结合千赫兹帧率两光子断层扫描(kilohertz-frame-rate imaging)和基于深度学习的神经网络(deep-learning-based neural network)实现了3D维度研究小鼠从出水口喝水时的舌部运动学。
首先,为了精准的从3D维度分解小鼠的舌部运动,研究者们在两个平面上以1kHz的频率观察记录舌部运动,并利用训练了两个深度人工神经网络来识别和区分小鼠的舌头。训练中,小鼠需要等待至少一秒以获得声音信号,然后在信号后1.3s内舔舐出水口就可以得到水分奖励(但小鼠是无法直接看到出水口的)。他们观察到小鼠的舌头在接触出水口之前和之后都有精细调控的可以分解的小动作,并将伸出舌头但还未找到并接触出水口前的小动作称为矫正子动作(corrective submovements,CSMs),而接触出水口后到缩回舌头之间的小动作称为“出水口接触子动作”(spout contact submovements,SSMs)。
CSM可以保证对点的准确性,它会非常有针对的往“记住”目标方向前进,并在此期间的加速峰值可以准确预测信号-与出水口接触的延迟。不仅如此,研究者们还注意到到在小鼠用舌头探索出水口期间(也就是第一次接触出水口之前的舌部运动尝试中),它们都出现的了非常明显的CSMs,而在找到出水口位置后,对CSMs的需求也就不那么大了。而如果增加实验间的间隔时长,小鼠的CSMs发生频率更高并且持续时间更长。所以说,CSMs在小鼠不确定目标位置的时候非常必须,在第一次舔到出水口确定了目标后,小鼠就会快速(大约找到目标0.1s)更新它们的动作计划。那么这个心机的小动作背后有着怎样不为人知的秘密呢?
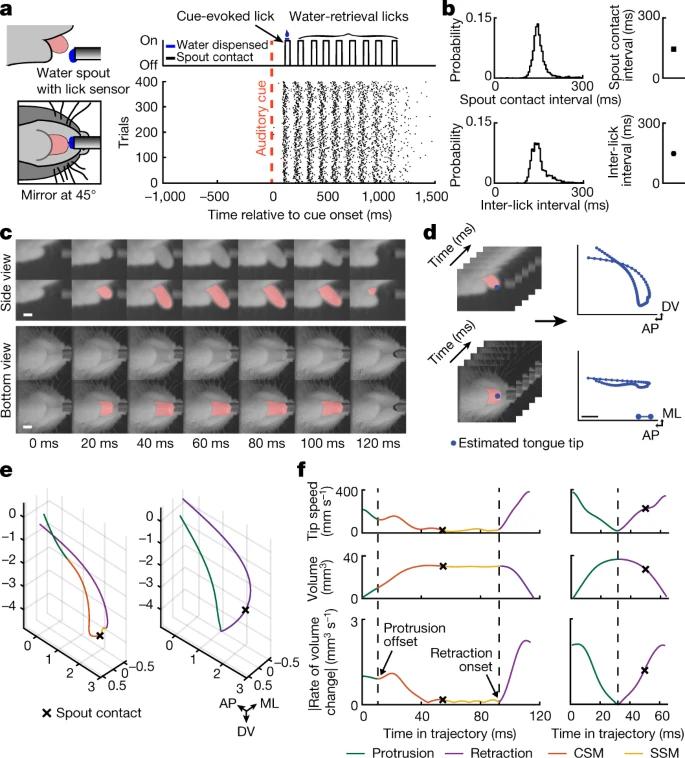 - Bollu et al., Nature -
- Bollu et al., Nature -研究人员猜想CSMs是不是也是受前外侧运动皮层(anterolateral motor cortex,ALM)控制的呢?他们发现用光遗传抑制ALM会部分损伤小鼠的反应动作(比如舌部探索时间更短,寻找路径更短速度更慢等)。并且经过ALM抑制后,小鼠则不会出现CSMs,而是探索方向错误时就立即缩回舌头,说明ALM的活动对舔舐动作的启动或瞄准并没有太大影响,而对纠错至关重要。
为了进一步确认ALM的神经元活动与CSM的关联,研究人员监控了ALM单个神经元的放电情况,发现早在CSM发生前ALM的神经元们就未雨绸缪,可以反应出正在进行的或者刚经历的CSMs。相反如果抑制ALM神经元,那CSM的执行以及“找到”路获取水分奖赏这一行为准确性都会大打折扣。这进一步说明了ALM的活动在CSM执行和监控期间的重要作用。
那么这是否也说明ALM的活动是我们更新信息纠错并调整行为的基础呢?接下来,研究人员设计了两步训练模式(double-step paradigm)来确认皮层在动作计划和执行修正的具体功能。这个任务可以检测小鼠是否可以整合一个舔舐动作内的所有探索信息然后进行动作修正。有趣的是,ALM正常活动时,小鼠可以非常迅速地判断更新后的出水口信息,并完成动作更正获得水源(比如通过加入CSMs或提前终止一次来回或增加探索距离等),而破坏ALM则严重损伤了小鼠的这种纠错能力。ALM的活动包含了所有发现出水口擦肩而过、产生CSMs、错过目标后的行动终止和找到目标确定位置等信息,是一个强大智能的CPU。
总的来说,这个研究首次综合利用了新的监测手段精细观察了小鼠舔水动作,并揭示了ALM对小鼠修正动作保证动作准确性过程的重要加持。如果你的舌头怎么都找不到你左前方那个可口的冰淇淋,放弃吧,可能是大脑皮层出了bug。
doi: 10.1038/s41586-021-03561-9
系统与网络
好奇心从何而来,又到何处去?
Ahmadlou et al., Science
@Veronica
乔布斯老爷子生前告诉过我们,做人要stay hungry,意即要始终保持求知若渴的好奇心。但说出来你可能不信,其实好奇心和吃喝拉撒睡一样,都属于动物的本能行为。很多时候,我们对新事物好奇也并不是为了追求它带来的奖励,有时候只是单纯地想了解而已。比如科学研究就向来如此,科学家们并不是奔着拿诺贝尔奖金去做研究的(当然如果有就最好~),他们只是跟随强烈的好奇心,一步步探索人类知识的边界。
小鼠也是一种好奇的生物,虽然它们眼神差,但碰到新东西也一定要闻闻摸摸,或许还会拽过来咬一口试试。一直以来科学家都认为小鼠的好奇心是一种捕食、觅食、吃东西行为(hunting,foraging,eating behavior),没什么值得大惊小怪的。但事实果真如此吗?最近来自伦敦大学学院的一项研究表明,小鼠的好奇心不同于捕食、觅食或吃东西,它们其实跟人类一样,对新事物有单纯的探索欲望,而这个欲望中心来自丘脑的未知核内侧(medial zona incerta,ZIm)。
科学家给小鼠设置了一个自由二选一测试(free-access double-choice test,FADC),让小鼠自由探索一个新物体或者一个熟悉物体。和预想的一样,小鼠对新物体的好奇心远大于熟悉物体,它们不仅会简单地接近和闻一闻(浅层探索),还会再进一步的拽一拽和咬一口(深层探索),而且小鼠对新物体的探索程度远比熟悉物体深入。接着,研究人员用钙信号成像技术检测小鼠在面对新物体时ZIm区域的神经元放电活性,他们发现,小鼠ZIm区域一群表达GAD2+的抑制性神经元(ZImGAD2)在探索新事物时放电程度大大提高了,而在它与食物互动的时候(此时小鼠的动作与探索行为相似)反应却很弱。如果用光遗传方法激活ZImGAD2神经元,则发现小鼠对新物体的探索行为和深度都显著增加。这些实验说明,ZImGAD2神经元能编码小鼠对新事物的探索行为。
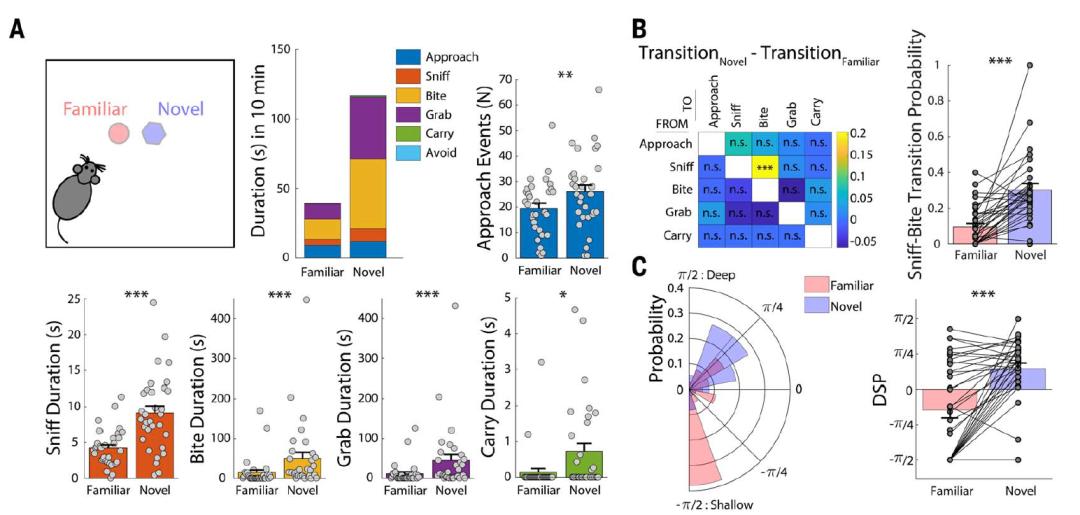 - Ahmadlou et al., Science -
- Ahmadlou et al., Science -由于ZIm在过去的研究中被报道与捕食有关,那么ZImGAD2激活到底是促进了对新物体的探索,还是单纯的促进了捕食呢?为了弄清楚这个问题,研究人员一口气做了四组FADC实验:1)两个物体的其中一个换成了熟悉的食物,另一个依然是新物体;2)两个物体的其中一个为熟悉的蟋蟀,另一个为与蟋蟀同步运动的新物体;3)两个物体的其中一个为活动的熟悉物体,另一个为静止的新物体;4)两个物体的其中一个为熟悉的蟋蟀尸体,另一个为静止的新物体。结果他们发现,一旦激活了ZImGAD2,无论是食物还是蟋蟀,管它是活的还是死的,只要是熟悉的物体都通通无法打动小鼠芳心了,它只对新物体情有独钟。值得注意的是,不仅仅是对新物体,小鼠对新伙伴的探索也同样受ZImGAD2控制,如果用光遗传技术分别激活或抑制ZImGAD2神经元,小鼠对新伙伴的探索行为和探索深度也相应的增加或减少了。这个结果除了证明小鼠是喜新厌旧的“花心大萝卜”之外,也意味着ZIm控制着小鼠对新事物、新伙伴的探索,而不是简单的捕食、觅食行为。
那么ZImGAD2上下游的调控通路各自为何呢?研究人员采用病毒示踪技术找到了ZIm的上下游投射区域。他们发现ZImGAD2的上游是前边缘皮质(prelimbic cortex,PL),如果激活PL→ZIm通路,则发现ZIm的放电频率变高了,觉醒水平也随之升高。而如果抑制掉PL→ZIm通路,则发现小鼠对事物和对小伙伴的探索行为都大大减少,也就是说,PL对ZIm可能释放了探索动力的信号,这对激发探索行为是至关重要的。不仅如此,研究人员还发现ZImGAD2的下游投射到了中脑导水管周围灰质侧面部分(lateral periaqueductal gray,lPAG),如果激活ZIm→lPAG通路,则会大大提高小鼠对新物体和小伙伴的探索行为。
最后,研究人员将ZIm抑制性神经元继续细分,试图找出具体哪一种抑制性神经元亚型导致了小鼠的探索行为。他们最终将目光锁定在表达tachykinin1(TAC1)的神经元上,这类神经元与其他抑制性神经元比如SST+、PV+并不同类。而且相比于ZI的头部和尾部区域,TAC1+神经元在ZIm区域表达量更高。果然,如果用光遗传技术特异性激活TAC1+细胞,就能大大增加小鼠对新物体、新伙伴的探索行为和深度,而激活其他抑制性神经元则没有明显反应。相反的如果抑制掉TAC1+神经元,小鼠的探索行为也被抑制了,这说明TAC1+神经元很可能是指挥小鼠探索行为的“幕后黑手”。
 - Ahmadlou et al., Science -
- Ahmadlou et al., Science -说来很有意思,zona incerta的拉丁文意思是“不确定的区域”,当时发现这个脑区的科学家Auguste Forel原话是“对于这个脑区,没有任何定论可说(a region of which nothing certain can be said)”。由于所知甚少,这个脑区当时甚至都没有被写进教科书。而今天本项研究告诉我们,ZI脑区恰恰是为了探究未知而存在的:一个不确定的区域负责探索未知,ZI真是恰如其名呀。
doi: 10.1126/science.abe9681
不同认知任务中
海马行波传递方向会反复横跳?
Kleen et al., Nature Communication
@Orange Soda
海马区在纵轴方向上与多个皮层和皮层下脑区连接,这种形态学连接可以帮助颞端(情绪信息处理相关)与大脑中隔端(视空间信息处理相关)功能连接,但大脑具体是如何实现这种多模态信息整合的,我们还不清楚。其中一种可能的机制就是:这一过程依赖于行波(traveling waves,TWs,在组织中传递的单个或周期性的电位变化),波的传递影响着神经活动的发生时间。此前在啮齿类和人类上的电生理结果报告了θ震荡(4-10Hz)从中隔端到颞端的传递;而与此同时有一些在大鼠上的研究则报告了源于海马的震荡传递可能并非沿着单一方向。另外,有研究表明朝着多个方向传递的震荡似乎与脑区之间不同种类的信息交互以及发放时间相关。因此Kleen等人想要探究人类海马震荡产生的行波是否会朝不同的方向传递,以及这其中可能表征着怎样不同的认知过程。
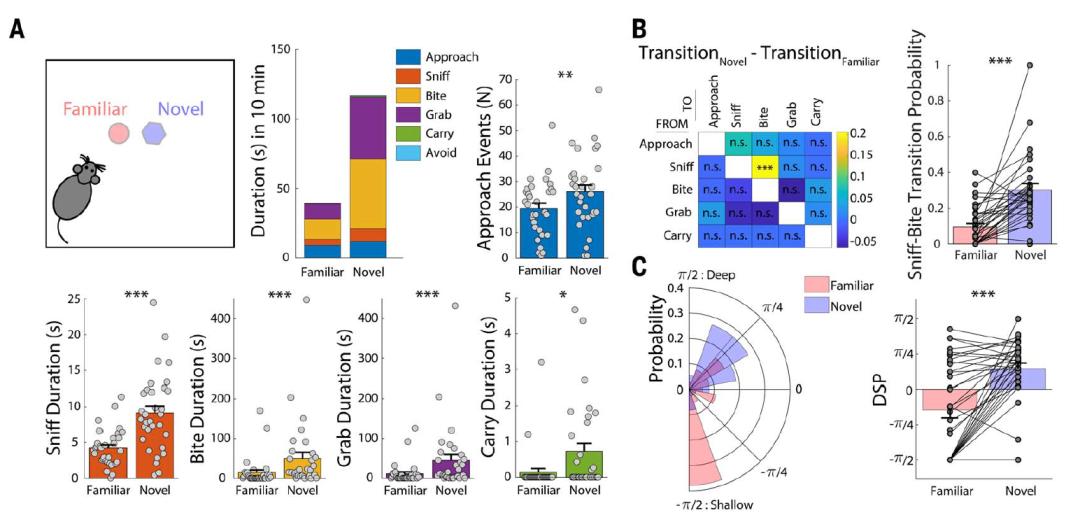
Kleen等人采用薄膜微电网阵列(thin-filmmicrogrid arrays;Fig.1a),将阵列放置在CA1区域附近(Fig.1b),这种新的技术让研究者可以在二维面上在体记录行波轨迹。一共6位被试(癫痫患者或脑部肿瘤移除手术过程中)参与实验,其中4位在手术中处于麻醉状态(右侧半脑),2位由于临床原因在术中处于清醒状态(左侧半脑)。
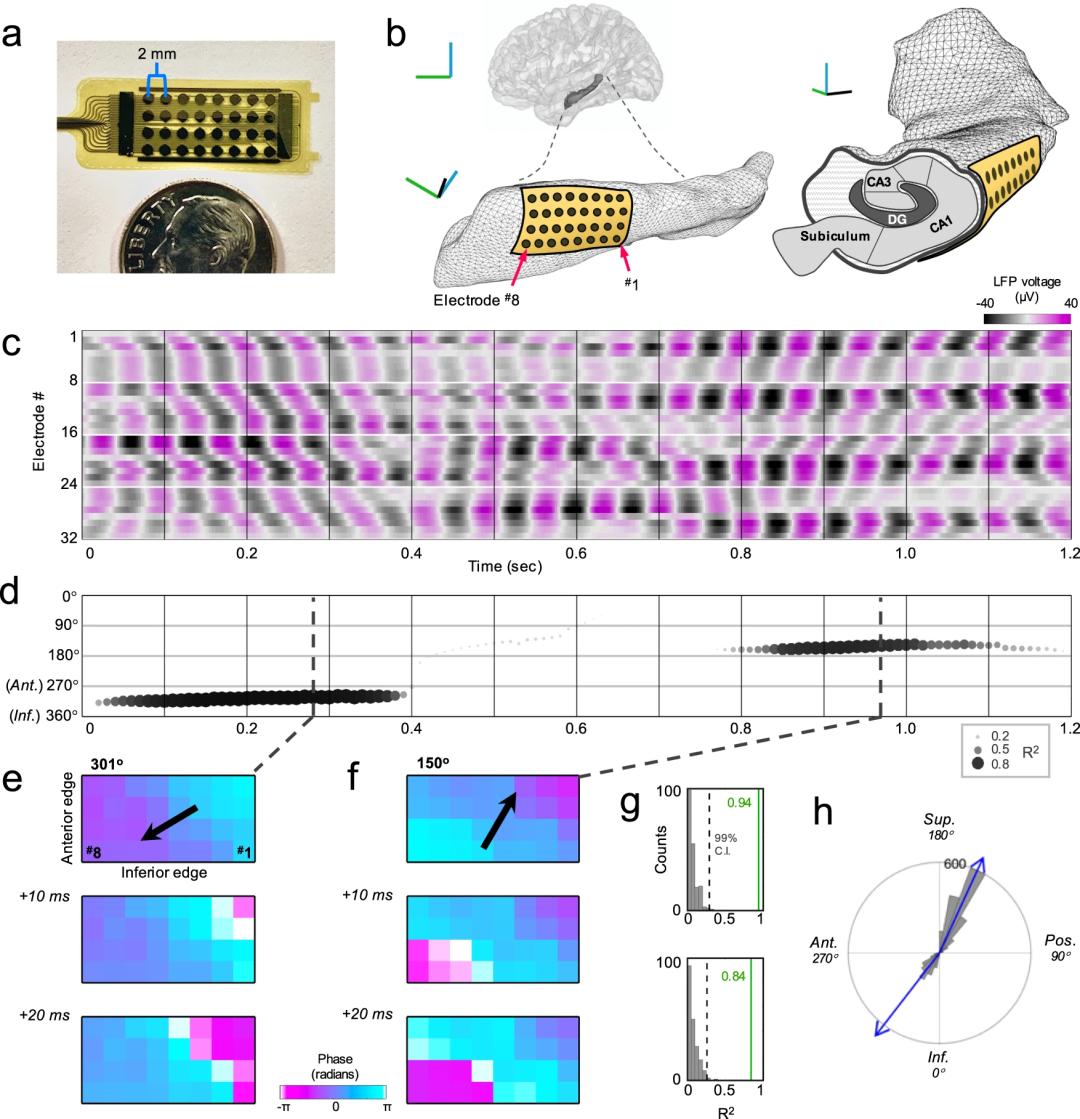 Fig. 1 | 在海马表面记录到朝两个不同方向传递的行波。
Fig. 1 | 在海马表面记录到朝两个不同方向传递的行波。—
Kleen et al., Nature Communication
对于采集到的局部场电位(local field potential,LFP)数据(1-15Hz),研究者首先寻找主导的频率峰,以这些峰为中心、带宽3Hz对数据进行带通滤波。如Fig.1c(示例的频率为13.8Hz)所示,相继电极所记录到数据表现出明显的相移。行波首先朝着向前向下(antero-inferior,AI)的方向传递几个周期后转变方向,朝着大约是相反的方向传播(postero-superior,PS)。并且统计发现在这一频段的行波几乎总是沿着这两个方向(虽然PS方向的传递更常见一些;Fig.1g,h)。Fig.2给出了所有被试的数据表现,在13位被试中有10位被试的数据表现出向两个不同方向传递的行波。另外,行波传递的方向在不同被试间高度一致,主要位于AI和PS两个象限(Fig.2b,e,f)。
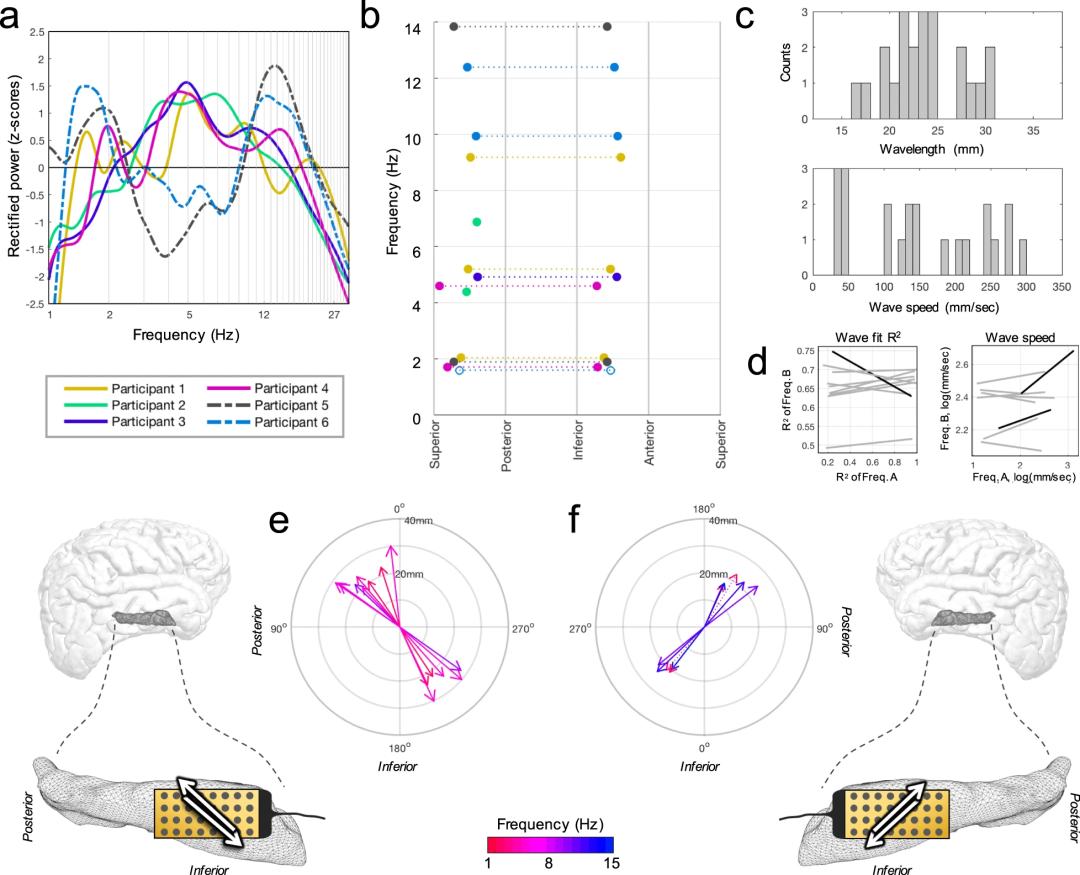 Fig. 2 | 所有被试的行波数据。
Fig. 2 | 所有被试的行波数据。—
Kleen et al., Nature Communication
由此Kleen等人的实验结果表明人类海马震荡产生的行波会朝两个不同的方向传递,那么不同方向传递的行波是否参与了不同的认知活动?
为了探讨这一问题,研究者接着在清醒被试上进行了视觉命名任务。实验中会呈现某个物体的图片,被试需要尽快说出该物体的名称。这项任务在手术中很容易进行,并有证据表明左侧海马会被激活。在这项任务中有两个任务相关事件:刺激出现和报告。研究者将数据与任务相关事件对齐后,计算行波传播方向的一致性(directionality consistency,DC)。发现大概在刺激出现后700ms,在1.9Hz频段朝着AI方向的DC出现了突然的提升(也就代表在AI方向上存在较为稳定的行波),这一现象在报告时也同样显著(Fig.3a,b),而在13.8Hz频段上的变化则并不明显(Fig.3g,h)。
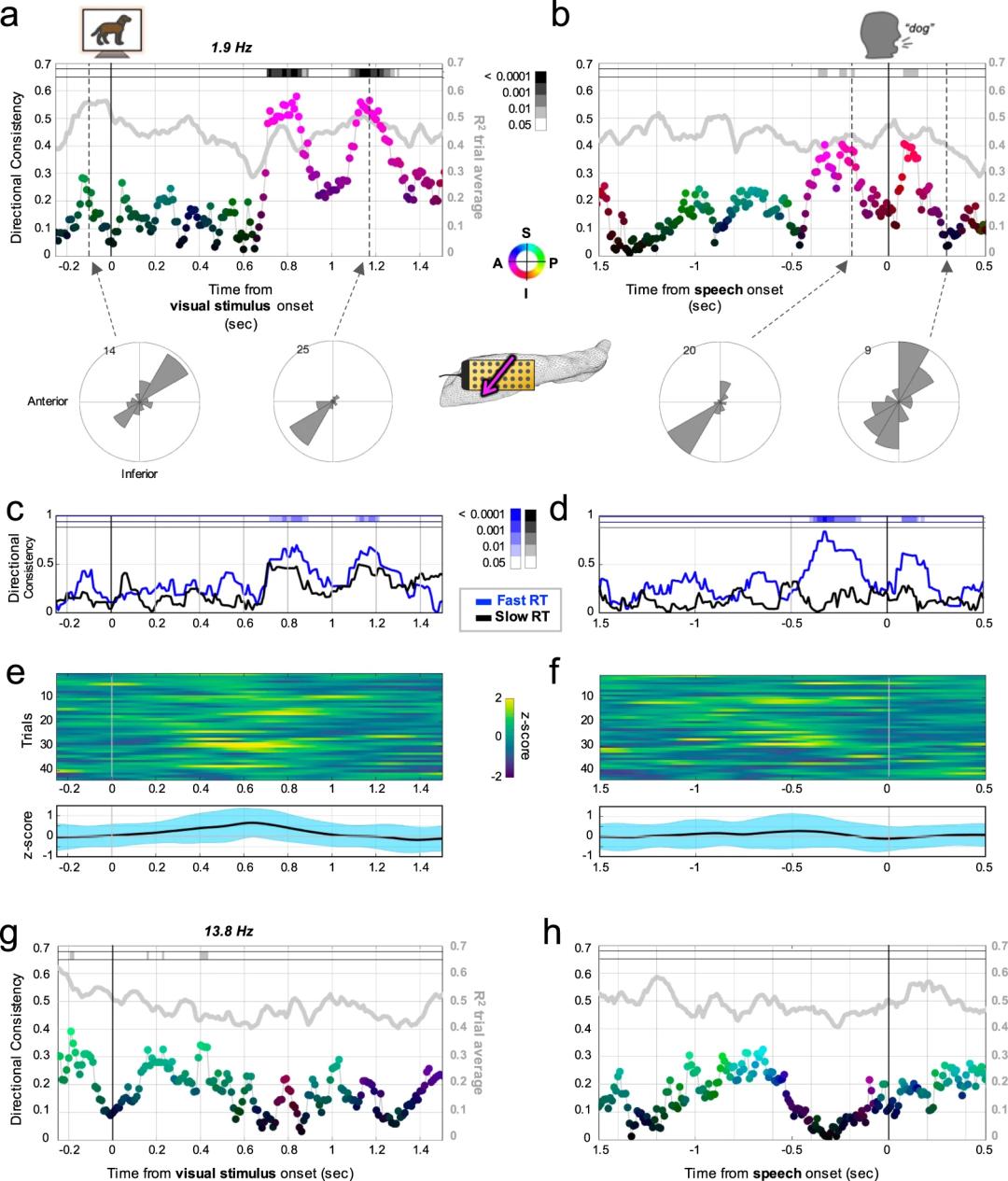 Fig. 3 | 视觉命名任务中的行波传递方向。
Fig. 3 | 视觉命名任务中的行波传递方向。—
Kleen et al., Nature Communication
研究者进一步分析了试次间没有任务的时间段(这一过程中被试处于期待阶段),发现在试次间1.9Hz和13.8Hz频段内都有朝着PS方向的DC值的提升(Fig.4),其中1.9Hz波段在试次-试次间的转换时表现出方向反转,而13.8Hz波段则会在试次间方向重置。
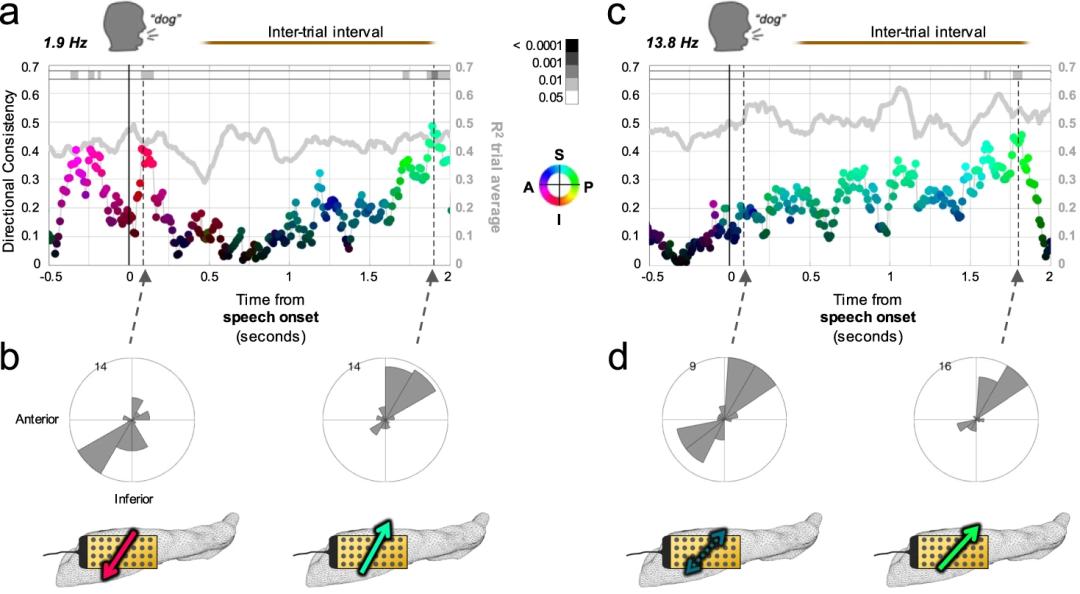 Fig. 4 | 试次间的行波传递方向。
Fig. 4 | 试次间的行波传递方向。—
Kleen et al., Nature Communication
根据在试次中(被试做任务阶段)和试次间(被试处于期待阶段)中行波传递方向的不同,Kleen等人推断,向两个不同方向传递的行波可能代表着在认知加工过程中不同的模式。具体来说,向前传递(anteriorly)的行波可能表示信息加工中预先更侧重于视空间信息的细节,接着再与情绪信息整合;相反,向后传递(posteriorly)的行波则预先更侧重于和视空间内容相关的情绪信息,然后再与具体的视空间内容联系起来。当前论文中的证据还较为初步,行波方向与认知过程之间的关系还需要更深入的研究来探讨。
doi: 10.1038/s41467-021-22850-5
编者:阿莫東森、Veronica、苏木弯、Orange Soda、图图
原标题:《熟人重要,还是族群重要?编码时空,我们和小鼠没有区别;阴差阳错,因未知命名的脑区竟字如其名 | Paper Alert # 29》