如何把气候塞进巫师的水晶球里?


你是否好奇过那些关于气候变化还有它们影响的预测是怎么做出来的?比如我们怎么知道未来地球会升温几度?会有哪些物种灭绝又会有哪些物种能够生存下来?这篇文章也许可以成为你理解这些问题的一个起点。

图源:Wikipedia
近几十年来,人类活动导致温室效应不断增强,引起了全球范围的气候变暖。这一趋势威胁着全球无数生物的生存。为了在变化的气候下保护全球生物多样性,并更全面地 评估未来气候变化在生态方面产生的影响,科学家已经提出多种分析气候变化对物种分布格局以及多样性的影响的方法论。 本文将重点关注其中知名度最高,应用程度最广的方法论之一: 生物气候包层模型(bioclimate envelope model),介绍其基本概念、分类标准、应用领域等方面。 虽然目前它面临着诸多方面的质疑,但在宏观尺度上充足的成功案例仍然足以肯定其在预测 气候变化对物种分布影响以及其他生态学研究领域的价值。
介绍
生物气候包层模型也被称为“生态位模型”,“栖息地适宜性模型”和“物种分布格局模型”。它通过寻找气候因素和物种在自然界中分布格局的联系,来定义一组物种可以在其中维持种群的气候环境条件。生物气候包层模型最常见的应用就是预测气候变化对物种分布格局的影响。
生物气候包层模型的根基来源于英国生态学及动物学家乔治·伊夫林·哈钦森于1957年提出的生态位理论(ecological niche theory)。哈钦森将基本生态位(fundamental ecological niche)定义为一个物种能够生存生长的 环境条件的集合。从定义上来说,“纯粹”的生物气候包层模型只将基本生态位中的气候变量因素纳入考虑,而不考虑其他可能影响物种分布的环境因素,如不同地区的土壤类型、资源丰富程度、以及捕食与被捕食的生物关系等。
基本生态位(fundamental ecological niche):基本生态位是一个物种能够生存的所有环境条件,由物种的变异和适应能力所决定,不受地理、捕食或者竞争等因素的影响。
实际生态位(realized ecological niche):基本生态位的一部分,考虑到了生物因素和它们之间的相互作用。或者说,是自然界中真实存在的生态位。
值得注意的是,虽然生物气候包层模型大部分的应用在于预测气候变化对物种分布格局的影响,但是在生态学的一些其他领域,比如自然保护区的地址选择和设计、评估未来物种入侵和疾病传播的危险、以及寻找适宜进行物种恢复、迁移或重引入的地区,该模型也可以为决策提供帮助。
两种广义分类标准及案例
气候变化包层模型可以按照两种广义分类标准而分成两大类。第一大类气候变化包层模型基于物种分布格局和环境变量之间的 实证关系,通过构建相关性模型预测气候对物种的影响。第二大类则基于生理学去分析气候参数与物种反应之间的机理。因此,这类模型是基于一个物种在气候层面上的生理限制来确定物种的气候生态位。

图源:DON GUTOSKI / WPY2015
基于实际生态位来预测气候对物种分布格局影响的相关性模型的一个早期应用便是Johnston所做的工作。他运用模型分析了入侵性多刺仙人掌的北美原产地的气候特点,从而预测了其在澳大利亚的入侵趋势。除此之外还有Hintikka的工作(引自Hengeveld, 1990),她参考地区性的温度变量,总结出一些欧洲物种正常活动范围内的气候条件,得以推测这些物种未来的可能分布。
相关性模型的另一个深入应用则是SPECIES模型。该模型借助人工神经网络(ANN),综合了观察到的物种分布格局以及五种气候变量。通过将该模型应用于一些欧洲高等植物物种,科学家们可以预测未来在气候变暖的情景下,这些物种可能的地理位置分布。
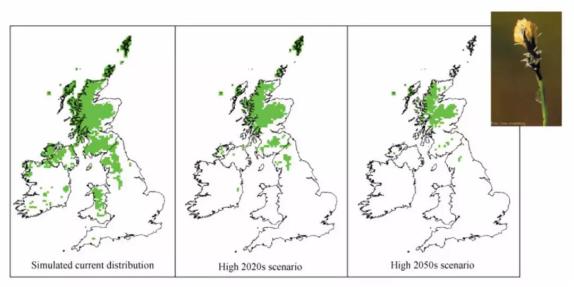
SPECIES模型基于英国未来气候场景模拟出的硬莎草重新分布格局。
来源:Pearson et al., 2002.
不同于基于实际生态位的相关性模型,生理学模型从潜在生态位角度研究气候对物种分布限制的机理。这种模型常常关注全球范围内生物群系(biome)的分布模式。例如,普伦蒂斯等人于1992年提出了一种预测全球生物群系的模型。该模型根据全球植物群系的生理数据,来计算每个植物群系所能生存的环境条件的极限。其预测出的全球植被分布模式与观测到的分布结果高度吻合。
生物群系(biome):生物群系是指适应某一特定环境类型(如沙漠,森林)的所有动植物群落的集合体。
所面临的质疑
尽管生物气候包层模型已经被运用于大量研究并作出了可靠的预测,近期的一些研究仍对其根本的正确性提出质疑,认为除了气候因素以外,许多其他因素对物种未来分布格局及动态也有着重要的作用。
质疑一
生物交互作用的影响
一些批评者指出,自然界中存在复杂的物种相互作用。单一物种分布的改变可能会影响其他物种的分布,但生物包层模型却忽视了物种间的 交互作用。例如,早在1961年,Connell就曾指出物种之间的竞争、捕食、共生的关系对未来物种分布格局的潜在影响。在他的一项关于藤壶(Chthamalus stellatus)在潮间带分布范围的限制因素的研究中,他发现该物种分布范围的下沿是由和另外一种藤壶(Balanus balanoides)之间的竞争关系以及和一种蜗牛(Thais lapillus)之间的被捕食关系所决定的。同样地,在Silander和Antonovics于1982年进行的盐沼地生态系统实验中,一种植物物种(Muhlenbergia capillaris)的消失会导致生态系统中其他五种植物分布范围同时扩大。如果不考虑生物交互作用,生物气候包层模型产生的预测就有可能严重偏离事实。

图片来源:missouri botanical garden
但包层模型的支持者认为,在宏观尺度上应用生物气候包层模型可以将生物交互作用的影响最小化。毋庸置疑的是,虽然在微观的地方尺度上,生物相互作用、地形、土地覆盖类型等可能会是决定物种分布格局的主要影响因素,但是在宏观尺度上,气候才是最重要的影响因素。事实上,许多生物气候包层模型在某些尺度上的应用非常成功地预测了物种当今的分布格局。例如Beerling等人于1995年应用气候响应面预测日本虎杖(Fallopia japonica)的分布格局,模拟情景与实际情境密切吻合的结果表明该物种在欧洲的分布的确是由气候因素主导决定的。

图片来源:Wikimedia Commons
质疑二
物种进化的影响
一些研究表明由气候变化引起的物种分布格局的变化不止是由于物种迁移到了新的适宜居住地带,同时也受到物种本身对环境做出的适应性进化的影响。Thomas等人在2001年便已验证过短期物种进化的重要性,他们重点关注了在过去20年间在英国扩大了地理分布范围的昆虫种类,发现有两种灌木蟋蟀 (Conocephalu sdiscolor和Metrioptera roeselii) 增加了长翅个体的比例,使得物种自身迁移能力更强;而两种蝴蝶(Hesperia comma 和Aricia agestis) 增加了它们可生存栖息地的多样性。因此,这些物种自身的变化都对生物气候包层模型的预测结果存在潜在影响。
然而,能够快速进化以应对气候变化的物种仍然是极少的。亚利桑那大学的Wiens等人曾研究了17个种群的570种现存脊椎动物,估计了每个种群相应地区到本世纪末的气候变化情况。尽管物种自身情况及所处气候有所差异,其结果却很一致:在17个种群中,15个种群的平均温度适应率慢于1℃/百万年;而两个稍微快些的种群也慢于2℃/百万年。这样的进化速度远远赶不上全球气候变暖的速度。2002年,在一项关于一种原生于美国大平原的豆类(Chamaecrista fasciculata)的研究中,埃特森和肖总结出这种植物预期的进化速率要远远慢于预期的气候变化速率。这些研究均表明,物种自身进化对生物气候包层模型的有效性所产生的影响很微小。

图源:Wikipedia
质疑三
物种迁移的影响
迁移能力强的物种可以通过不断迁移来追踪适宜它们生存的地理环境。物种的迁移能力不仅取决于其自身的生理特征,还取决于迁移发生的 地形:包括“天然屏障”-如山脉地形,和人为导致的栖息地分裂和破坏-如森林砍伐、城市化。因此,要准确预测物种未来的分布格局就需要详细了解该物种在其不断变化的地理情况以及气候因素限制下的迁移能力。因此,生物气候包层模型所作出的预测很可能与实际结果不符。
然而古生态学记录中的一些证据表明气候上的变化足以解释植物在大陆性尺度下的迁移。事实上,近期的研究也强调了物种借助于罕见的长距离迁移来应对气候变化的能力。这一观点在从古生态学到现代的外来植物物种传播的观察中都得到了例证。Johnson和Webb于1989年也提供了支持生物气候包层模型的证据,他们发现北美东部一种壳斗科植物迁移速率的差别主要源自于个体对于气候变化反应。因此,在合适的尺度上应用生物气候包层模型仍然可以提供较为准确的预测。
总结与展望
考虑到自然系统的复杂性,生物气候包层模型所预测的结果会不可避免地与实际情况存在或大或小的偏差。所以,我们应以谨慎保守的态度来看待生物气候包层模型所作出的预测,把它视为体现气候变化对未来物种分布格局及其他诸多方面潜在巨大影响的近似估计,而非准确的预测和模拟。但是我们仍应肯定生物气候包层模型的作用,有了它提供的较为准确的结果,人们就能够更好地理解复杂的环境因素对物种的潜在影响,从而指导未来的野生动物保护。

图源:网络
想要更真实准确地模拟气候变化对物种分布的影响,我们需要更好地理解影响物种分布格局的诸多因素之间复杂的相互作用,并根据不同情况选择最适宜的模型和技术。
本文由前三篇文献概译
Pearson, R. G., & Dawson, T. P. (2003). Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? GlobalEcology & Biogeography (Vol. 12).
Jo, M. B. A., & Peterson, A. A. T. (2012). Uses and misuses of bioclimatic envelope modelling. Ecology, 93(7), 1527–1539.
Heikkinen, R. K., Luoto, M., Araújo, M. B., Virkkala, R., Thuiller, W., & Sykes, M. T.(2006, December). Methods and uncertainties in bioclimatic envelope modelling under climate change.
参考文献:
Johnston, T. H. (1924). The relation of climate to the spread of prickly pear. Royal Society of South Australia.
Hengeveld, R. (1992). Dynamic biogeography. Cambridge University Press.
Pearson, R. G., Dawson, T. P., Berry, P. M., & Harrison, P. A. (2002). SPECIES: a spatial evaluation of climate impact on the envelope of species. Ecological Modelling, 154(3), 289-300.
Woodward, F. I., & Woodward, F. I. (1987). Climate and plant distribution. Cambridge University Press.
Prentice, I. C., Cramer, W., Harrison, S. P., Leemans, R., Monserud, R. A., & Solomon, A. M. (1992). Special paper: a global biome model based on plant physiology and dominance, soil properties and climate. Journal of biogeography, 117-134.
Connell, J. H. (1961). The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology, 42(4), 710-723.
Silander, J. A., & Antonovics, J. (1982).Analysis of interspecific interactions in a coastal plant community—a perturbation approach. Nature, 298(5874), 557.
Beerling, D. J., Huntley, B., & Bailey, J.P. (1995). Climate and the distribution of Fallopia japonica: use of an introduced species to test the predictive capacity of response surfaces. Journal of Vegetation Science, 6(2), 269-282.
Davis, M. B., & Shaw, R. G. (2001). Range shifts and adaptive responses to Quaternary climate change. Science, 292(5517), 673-679.
Thomas, C. D., Bodsworth, E. J., Wilson, R. J., Simmons, A. D., Davies, Z. G., Musche, M., & Conradt, L. (2001). Ecological and evolutionary processes at expanding range margins. Nature, 411(6837), 577.
Graham, R. W., & Grimm, E. C. (1990).Effects of global climate change on the patterns of terrestrial biological communities. Trends in Ecology &Evolution, 5(9), 289-292.
Collingham, Y. C., Hill, M. O., & Huntley, B. (1996). The migration of sessile organisms: a simulation model with measurable parameters. Journal of Vegetation Science, 7(6), 831-846.
Clark, J. S., Fastie, C., Hurtt, G., Jackson, S. T., Johnson, C., King, G. A., ... & Schupp, E. W. (1998). Reid's paradox of rapid plant migration: dispersal theory and interpretation of paleoecological records. BioScience, 48(1), 13-24.
Higgins, S. I., & Richardson, D. M. (1999).Predicting plant migration rates in a changing world: the role of long-distance dispersal. The American Naturalist, 153(5), 464-475.
Cain, M. L., Milligan, B. G., & Strand, A.E. (2000). Long‐distance seed dispersal in plant populations. American journal of botany, 87(9), 1217-1227.
Johnson, W. C., & Webb III, T. (1989). The role of blue jays (Cyanocitta cristata L.) in the postglacial dispersal of fagaceous trees in eastern North America. Journal of Biogeography, 561-571.
编译 | 孙祥荣
小土豆班成员。来自北京的高中生,对气候变化,可持续发展和野生动物保护等领域十分着迷;同时十分热爱金属乐和电吉他。
编辑 | 王柳枫